

Hello! We are the James Lab for X-SIG 2024. Here, we use the filamentous fungus Aspergillus nidulans as our model organism to uncover the intricate molecular processes underpinning eukaryotic cellular function. We use genetic engineering to directly modify a gene-of-interest. This might entail attaching Green Fluorescent Protein (GFP) to a gene so that we may track the intracellular localization of a protein with a fluorescence microscope; adding a biochemical tag (e.g., HA3) so that we might detect binding partners of the encoded protein; or deleting the gene outright so that we might observe the phenotype that results from our meddling.

These images are of A. nidulans, and the web-like structures are the filaments, termed hyphae. This characteristic feature of A. nidulans designates it as a filamentous fungus.
At present in the James Lab, we are expending most of our energy attempting to understand the function of a novel (new-to-science) gene that Dr. James discovered, called wmcA. This gene is of particular interest because it is a major regulator of the cytoskeleton, which provides structure, support, and organization for the cell. There are a number of different structures associated with the cytoskeleton, but wmcA specifically governs the function of one of three major players–microtubules (MTs). MTs play a critical role in structure, cargo trafficking, and mitosis. Thus, genes that encode proteins that significantly affect the function of MTs have great and far-reaching consequences for the cell and are of great interest due to such importance.
Specifically, when the wmcA gene is deleted from the genome of A. nidulans and the fungus is incubated at low temperatures (~22 °C), they undergo a dramatically named phenomenon known as mitotic catastrophe, which is defined as the absence of MTs and the dispersal and fragmentation of chromosomes. This tells us that wmcA is essential to MT homeostasis and correct cellular function. The severity of the consequence for cells without wmcA bespeaks its importance.
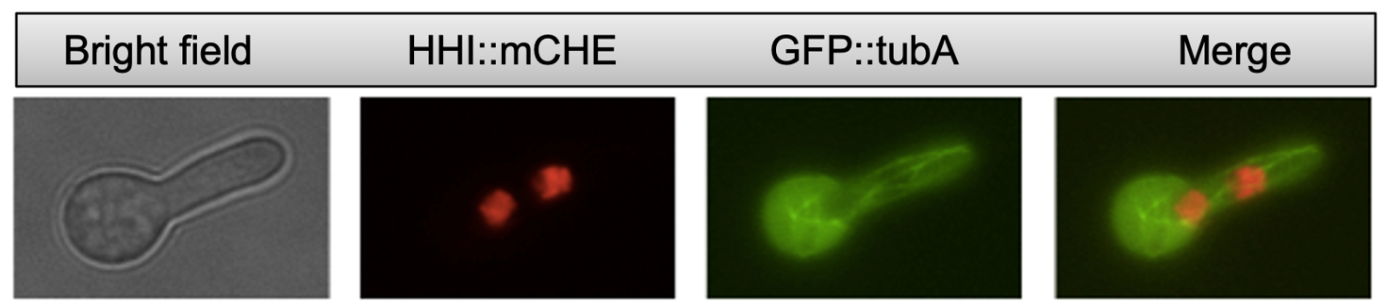
This is an image taken under the fluorescence microscope, in which GFP has been added to a building-block of MTs, called tubulin. This gene construct (GFP::tubA) was made via PCR and incorporated into the genome of A. nidulans through a process known as transformation. The green, string-like structures are MTs within a live cell.
What is more, the gene wmcA is specific to filamentous fungi. A number of species of filamentous fungi are pathogenic, to both humans and plants. One such fungus, Magnaporthe oryzae, also known as the rice blast fungus, has a detrimental impact on agriculture, destroying one-third of the total rice harvest per annum. Given that rice is the primary staple food for more than half the world’s population (USDA), M. oryzae infection poses a threat to the majority of the world’s food supply. As such, wmcA may be a valuable antifungal drug target.
First, we need to further elucidate the function and the specific cellular pathways in which wmcA is involved. Part and parcel of understanding the function of a gene is to investigate other genes with which one’s gene-of-interest interacts. This approach gives clues as to the different pathways and mechanisms by which a gene acts. In previous years, the James Lab discovered a number of proteins that interact with wmcA, which attest to the scope and importance of wmcA. Moreover, uncovering these genetic interactions also expands our understanding of the genes that interact with wmcA. In all, the James Lab has accrued an increasing number of genes to investigate along our journey to gain a clearer picture of the molecular pathways in which wmcA is involved. See “Specific Projects” below for some more information about the genes that interact with wmcA.
Meet the Lab!

From left to right: Merlin, Dr. James, Alessandro, and Sarah. These awesome shirts were hand-crafted by Sarah using the print shop on campus.
Alessandro
Hey, I’m Alessandro. I am currently a rising senior, majoring in Biochemistry & Molecular Biology with a minor in History. I joined Dr. James’s lab in the Spring 2024 semester and have continued my work into the summer. So far, most of my work has centered around mcnC and exploring its functions. I plan to continue my work with Dr. James throughout the entirety of my senior year.
Sarah
Hi! 🙂 I’m Sarah, a rising senior with a self-designed major in Forensic Chemistry. I have been a part of Dr. James’ lab since the Fall 2023 semester and plan to continue until graduation. Along with studying benA in aspergillus, I will also be starting my senior capstone project of identifying the (many…many) unknown fungal contaminants of the Science Center. If your lab is contaminated, please (please) give it to me. Don’t worry, we won’t create “The Last of Us” this summer. Probably.

Merlin
Warm greetings from your apprentice wizard! I’m a rising junior of Biochemistry & Molecular Biology with great binding affinity toward molecular genetics and bioorganic chemistry. I have been with my maestro and peers since this Spring to rescue my favorite food, rice, from the blast fungus. I’m investigating the interactions between 2 proteins, TBCA and wmcA, in A.nidulans. My hobbies include hiking at the Appalachian Trail, playing with cats (cat dad 24/7), and making fun mnemonics for Biochemistry (for next semester, as below).

Specific Projects
Below are the four main genes of interest that we in the lab are investigating due to their suggested link to wmcA function.
mcnC
The James Lab discovered that a gene named mcnC has a genetic interaction with wmcA. That is to say that manipulation of both genes–mcnC and wmcA–within a single cell leads to a unique and interesting phenotype. Recall that cells in which wmcA had been deleted were characterized by mitotic catastrophe. However, when cells that already had a wmcA deletion had deleterious mutations in the mcnC gene, the cells grew much better at lower temperatures. This may seem somewhat surprising at first, but this suggests that mcnC works in opposition to wmcA. That is, while wmcA promotes MT homeostasis, mcnC somehow functions the opposite way–but we don’t know how. Just like the government, there must be checks and balances within the cell, so having genes that work in different directions ensures that homeostasis is maintained.
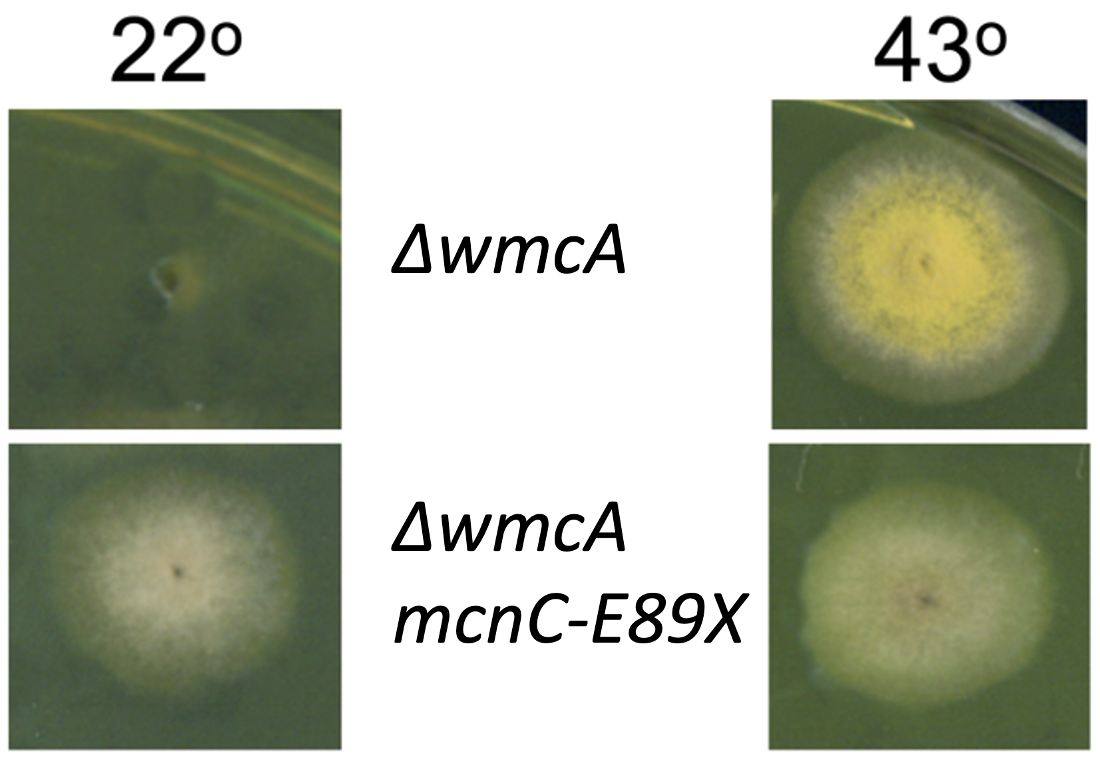
At 22 °C, cells with a deletion of wmcA (∆ = deletion) cannot grow significantly. However, combining ∆wmcA with a mutation that negatively affects mcnC function (e.g., mcnC-E89X) rescues the cell at this lower temperature, and A. nidulans is able to grow.
Although we might be a bit biased, we think mcnC is a very interesting gene. Its implication in MT stability via its genetic interaction with wmcA is new-to-science. Previous work that initially identified mcnC found that it was involved in the cell cycle and identified a special domain within the encoded protein called a CUE domain. This allows mcnC to bind to ubiquitin, which is a widespread marker for degradation of proteins. Hence, mcnC might regulate other proteins by promoting their destruction. In fact, the ortholog (a gene that performs a similar function but in a different species) in Saccharomyces cerevisiae, or yeast, is called Def1 and is involved in the degradation of a protein that makes RNA from DNA in a process called transcription. This suggests that mcnC in A. nidulans might be involved in ubiquitin-mediated degradation of wmcA. To test this hypothesis, we plan to perform a coimmunoprecipitation assay (CO-IP) to see if mcnC and wmcA physically interact. We also plan to see what part of mcnC is essential for its function, as evidence suggests that the entirety of the sequence of the encoded protein is not necessarily essential.
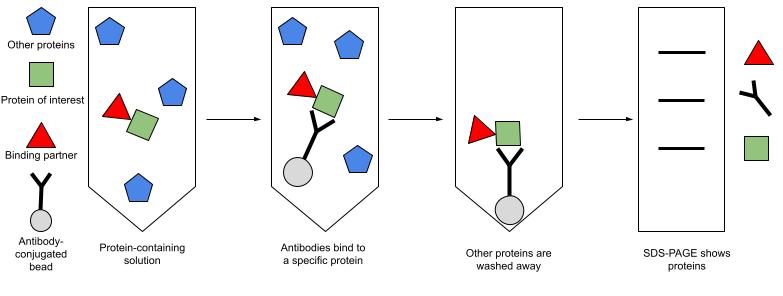
A CO-IP uses antibodies–used for their marked ability to bind to one specific protein–to “capture” a protein of interest. If two proteins physically associate by binding to one another, then both proteins should be “captured.” We can then identify any protein binding partners.
VTS1
VTS1 is a developmental regulator and another interesting gene. Recently, phosphorylation of the encoded VTS1 protein was found to be required for the virulence of M. oryzae, the rice blast fungus. Only a few weeks ago, Dr. James was notified that VTS1 physically interacts with mcnC in M. oryzae. VTS1 is also found in A. nidulans, so this evidence suggests a novel function for mcnC. Could mcnC be marking VTS1 for destruction via ubiquitination? Or is mcnC performing another function?
benA
Microtubules are entirely composed of tubulin, a heterodimer which contains 𝛂 and 𝛃-tubulin. BenA is one of two genes that encodes for 𝛃-tubulin. Overexpression of beta-tubulin has been shown to lead to a loss of microtubule structures and a loss of cell viability. My project for the summer is to figure out if wmcA is physically associated with benA. So far, we have found that molecular disruption of benA via a GFP tag at the C terminus results in strong growth impairment, meaning that GFP tag may disupt 𝛃-tubulin function. Future experiments include the manipulation of temperature in mutant benA and wmcA strains to observe microtubule growth.
TBCA
The newly synthesized 𝛃-tubulin polypeptide must be folded into a proper 3D structure before functioning. This process is uniquely assisted by TBCA (Tubulin Binding Cofactor A). Without functional TBCA protein, unfolded 𝛃-tubulin cannot polymerize to form the microtubule; the fungal cell fails to reproduce and becomes nonviable. As common sense, we would expect the double mutant of both wmcA and TBCA (ΔTBCA ΔwmcA) to exhibit an even worse growth than the single mutant wmcA. Surprisingly, our genetic data showed the opposite: the double mutant is living more happily than its counterpart. In other words, the mutation of ΔwmcA miraculously rescued TBCA mutant cells from disaster! This intriguing paradox is where our journey with TBCA begins.
It turns out the world of proteins is dynamic: proteins can physically bind, destroy, inactivate, or activate one another. With TBCA and wmcA, we are on an endeavor to discover the relevant interactions between them. By computational work (in silico) with ChimeraX, my maestro has found suggestive evidence of physical binding between this pair. My plan for the summer will be to confirm if this is also true with in vivo work with A.nidulans. This work would support more understanding about the genetic and molecular roles of wmcA in microtubule stabilization.
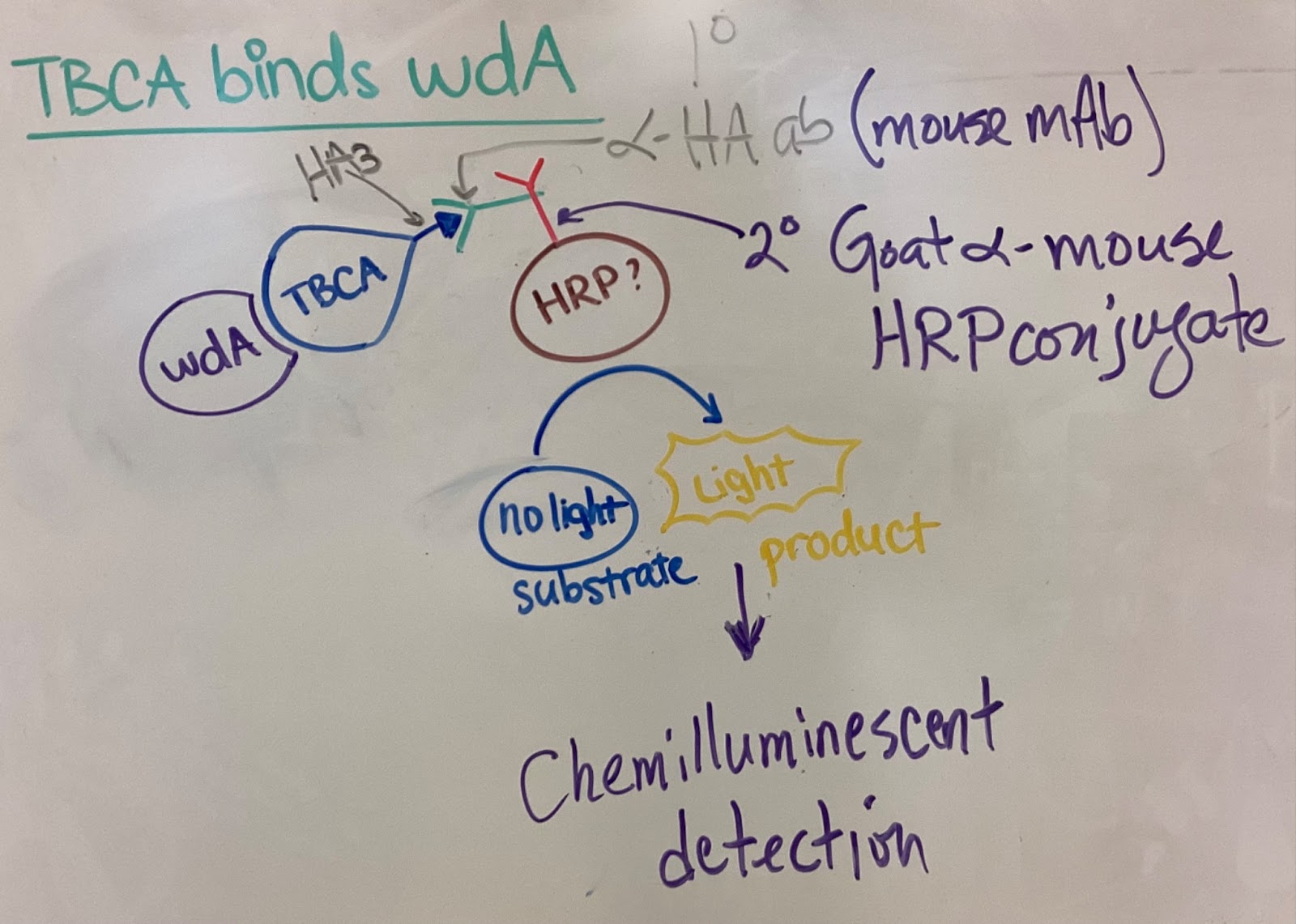
For my project, TBCA protein is not attached with a big fluorescence tag (which might disrupt its potential physical interactions with wmcA), but instead a small epitope tag called HA3 is tagged for visualization (TBCA::HA3). A GFP-tagged allele of wmcA (wmcA::GFP) is isolated with a highly specific anti-GFP antibody (this technique is called immunoprecipitation). With proper techniques, we preserve relevant protein-protein physical interactions, “pulling out” complexes of wmcA and its binding proteins. These complexes are separated by molecular weight with Western blot and incubated with anti-HA3 primary antibodies. A secondary antibody, conjugated with horseradish peroxidase (HRP), binds specifically to the primary antibody. HRP can convert a substrate that we use from non-illuminating to illuminating, which can be visualized under UV. Overall, if TBCA binds to wmcA, this chemiluminescence is observed and vice versa. The principle of this visualization is shown below.
Thanks for reading! 🙂 – The fun-guy (and gal) lab
